|
Circadian rhythms in cyanobacteria
The core oscillator of Synechococcus elongatus is encoded by the kaiA, kaiB and kaiC genes, which regulate global patterns of gene expression and the timing of cell division. KaiC is an autokinase, autophosphatase, and ATPase, whose daily rhythms of phosphorylation are key features of the timekeeping mechanism that drives circadian rhythms in complex with KaiA and KaiB. These endogenously generated rhythms are synchronized with the environment through an input pathway that monitors the cellular redox state. Temporal information from the clock is related through an output pathway consisting of histidine kinase SasA and cognate response regulator RpaA, and a phosphatase, CikA that dephosphorylates RpaA. We seek to understand the molecular mechanisms by which the cyanobacterial circadian clock functions in the context of the three-dimensional cell. We use a variety of techniques including cell and molecular biology as well as genetics and biochemistry to understand what cellular factors the clock interacts with in order to in order to synchronize with its external environment and execute programs of cellular physiology. Cell biology of the clock We found that core oscillator component KaiC localizes as discrete foci near a single pole of cells in a clock-dependent fashion, with enhanced polar localization at night. Current projects are aimed at understanding the molecular mechanism by which KaiC localizes to the pole at night and elucidating the spatial distribution of the clock network components in order to determine how subcellular localization of clock components and the co-localization of factors that vary in space and time contribute to the clock system. Roles for protein degradation in regulating the circadian clock Protein degradation is an important quality control mechanism necessary for a broad range of cellular processes, including the circadian clock. ClpXP mediated proteolysis plays critical roles in controlling cell cycle progression, establishing cell polarity, competence and sporulation in other bacterial systems. The S. elongatus genome encodes one homolog of ClpX, an ATPase required for substrate recognition, and three ClpP peptidase paralogs. ClpX also possesses protein remodeling functions, independent of ClpP, where ClpX-induced structural changes of protein substrates results in altered biological activity. Roles for the clpXP gene products have been reported in both cell division and circadian rhythms: clpX mutants display elongated cells, and disruption of clpX, clpP1 or clpP2 results in long-period phenotypes. Our results suggest that while ClpXP protease functions are required to maintain normal periodicity of the circadian clock and the clocks ability to synchronize with the environment, remodeling functions of ClpX are required for circadian control of cell division. We seek to elucidate the proteome changes that are occurring that regulate cells ability to synchronize with the external environment. Additionally we aim to understand the molecular details of how ClpX regulates circadian timing of cell division. |

Our model organism Synecococcus elongatus PCC7942.
The core oscillator of S. elongatus consists of KaiA, KaiB and KaiC.
Image courtesy of the UCSD BioClock Studio
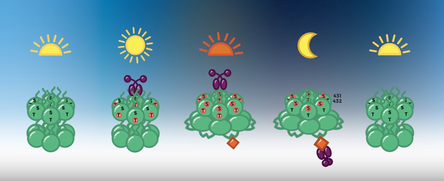
KaiA association with KaiC during the day promotes KaiC's autophosphorylation on neighboring Thr and Ser residues. Once KaiC is fully phosphorylated a binding site for KaiB on KaiC is exposed. KaiB can then bind to KaiC and sequester KaiA in an inactive form, promoting KaiC auto dephosphoryaltion at night. Image courtesy of the UCSD BioClock Studio.

Fluorescent micrograph showing KaiC (visualized using a YFP-KaiC translational fusion in green) localization in the cell (cell membrane shown in red). During the day KaiC is diffuse throughout the cell and at night KaiC localizes to one cell pole.
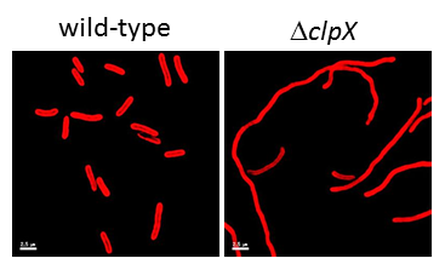
clpX mutant cells (right panel) have an elongated cell morphology compared to wild-type cells (left panel) suggesting that they have a defect in cell division. Autofluorescence from the photosynthetic pigments in teh membrane outline the cells in red.
|
